La continuidad cíclica de los seres vivos encuentra los vínculos entre generaciones sucesivas en los fenómenos de reproducción.
La reproducción se realiza a distintos niveles de la escala evolutiva, en las distintas ramificaciones de los reinos vegetal y animal, en las distintas especies vivientes, con una variedad de mecanismos que por sí sola justifica todo un tratado.
Una primera clasificación de los fenómenos de reproducción debe distinguir los organismos unicelulares de los multicelulares, ya que sólo en los primeros coincide la división celular con la reproducción.
En multicelular, la reproducción puede ser agámica o sexual (o gámica).
La reproducción agámica, relativamente menos frecuente, se basa en el mecanismo de la mitosis, por lo que la variabilidad de las especies se confía más bien a la aparición repetida de mutaciones.
También existen diferentes mecanismos, como la estrobilización, la esporulación, etc., mientras que en las plantas encontramos formas regenerativas muy conocidas en la agricultura (esquejes, acodo, etc.).
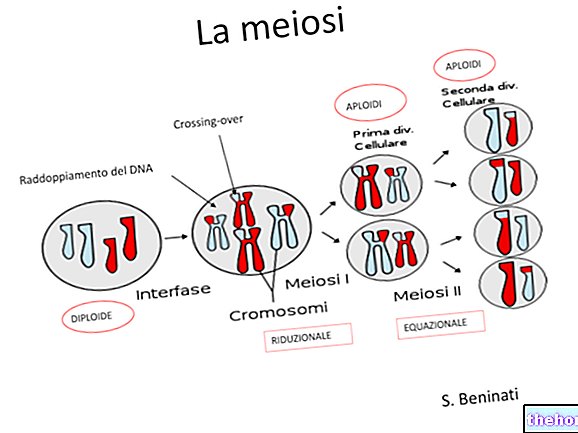
El mecanismo de reproducción más extendido en las formas superiores, sin embargo, es el sexual, correspondiente a la aparición de la meiosis, la formación de gametos y su fusión en el cigoto (fecundación).
En las especies primitivas los gametos no están diferenciados morfológicamente: en este caso hablamos de isogametia. Sin embargo, siempre hay dos series de gametos, identificados con los símbolos (+) y (-) y la fecundación sólo puede tener lugar a partir del "encuentro entre gametos de signo opuesto: hay por tanto una diferencia biológica, aún no expresada morfológicamente".
A medida que avanza la escala evolutiva, aparece una diferenciación morfológica y funcional, en la que un tipo de gametos femeninos generalmente dotados de abundante material de reserva (el deutoplasma o ternero, que asegurará el desarrollo del embrión hasta que pueda ser metabólicamente independiente) y un tipo de Los gametos masculinos dotados de motilidad para llegar a los femeninos, siempre son haploides y son el resultado de la meiosis, su fusión da lugar al cigoto diploide.
Entre la meiosis y el cigoto pueden transcurrir una serie de generaciones de células haploides, así como entre el cigoto y la meiosis pueden transcurrir una serie de generaciones de células diploides, con las diferentes variedades de ciclos generativos expresados por la alternancia de generaciones.
Una especie diplonte (con un organismo diploide) se caracteriza por la meiosis gamética: la meiosis produce directamente los gametos, que al fusionarse inmediatamente reconstituyen el estado diploide. Este es el caso predominante de los metazoos, incluido el hombre.
Gametogénesis
Teniendo en cuenta la reproducción con meiosis gamética, como ocurre en el hombre, intentemos aclarar cómo encaja la meiosis en la gametogénesis (formación de gametos).
En el desarrollo embrionario, de gametogénesis masculina y femenina (llamado espermatogénesis y ovogénesis), existe una diferenciación temprana entre las células destinadas a formar el cuerpo (línea somática) y las destinadas a producir gametos (línea germinal). Las células iniciales de la línea germinal se denominan protogones. Con la diferenciación de la gónada en el sentido masculino o femenino, se produce la diferenciación de las células germinales, respectivamente, en espermatogonias y ovogonias.
Al observar la espermatogénesis, vemos que hay una serie de generaciones de células en las espermatogonias, que continúa durante toda la vida. Sólo una parte de las espermatogonias así producidas continuamente difiere del ciclo mitótico normal y en su lugar comienza el meiótico.
La célula germinal en la que comenzará la meiosis (reduplicación y luego primera división) se llama espermatocito de primer orden; su división da lugar a dos espermatocitos de segundo orden, que con la segunda división dan lugar a un total de cuatro espermátidas.
Podemos graduar la reducción del kit cromosómico de 4n de los espermatocitos de primer orden (después de la reduplicación hay cuatro cromátiles para cada par de homólogos) al 2n de los espermatocitos de segundo orden y al n de las espermátidas, como ya se vio al estudiar meiosis, que Así concluimos. Las espermátidas ya son haploides, pero aún no son gametos maduros. A partir de la estructura de una célula de tipo haploide, la maduración funcional (llamada espermiohistogénesis) transforma las espermátidas en espermatozoides, es decir, gametos masculinos maduros.
En la gametogénesis femenina (u ovogénesis) existen varias diferencias. En primer lugar, el número de gametos a preparar es mucho menor. Se estima que en las gónadas de una hembra de la especie humana se preparan alrededor de 5 x 105 ovogonos; de estos, solo alrededor de 400 están interesados en la maduración del folículo y el posterior abatimiento, en un ciclo que generalmente afecta solo un folículo por mes durante un período fértil de aproximadamente 35 años.
El diferente número de gametos preparados en los dos sexos corresponde a la diferencia de función y comportamiento ya mencionada: los espermatozoides son pequeños, móviles y numerosos en relación a la necesidad de buscar el óvulo y la baja probabilidad de encontrarlo; los óvulos son grandes, inertes y escasos, en relación a la función de garantizar al embrión el material de reserva y la protección que le proporciona la fecundación interna (naturalmente, sobre todo en la fecundación externa los óvulos también deben ser más numerosos).
La necesidad de dotar a los gametos de material de reserva corresponde a la presencia, en la "ovogénesis, de una fase de detención de la meiosis, durante la cual los cromosomas se desespiralizan parcialmente. A continuación, se observan los denominados" cromosomas pluma ", en los que una serie de extraversiones identifica los rasgos en los que se desreprimen los genes responsables de la síntesis de deutoplasma.
El menor número requerido para los gametos femeninos también corresponde al hecho de que, de las cuatro células haploides producidas por la meiosis, solo una recibe todo el material de reserva y se convierte en gameto, mientras que las otras tres (los polocitos o cuerpos polares), que contienen solo material cromosómico, no pueden dar lugar a cigotos y embriones y están destinados a retroceder.
Fertilización
La fertilización, es decir, el encuentro entre el gameto masculino y femenino, se puede realizar de formas muy distintas.En el reino animal observamos el paso de la fecundación externa (gametos expuestos a cualquier riesgo ambiental y por tanto necesariamente muy numerosos en los dos sexos) a la fecundación interna, a la que el cuidado parental está además ligado a la relación metabólica de los mamíferos entre madre y feto. .
La fertilización, una vez que se ha producido el encuentro entre gametos del sexo opuesto, debe tener lugar garantizando dos condiciones: especificidad y unicidad. Es decir, se debe asegurar que el espermatozoide sea de la misma especie que el óvulo y que, una vez que entre el primero, no entren otros.
La especificidad está asegurada por las características bioquímicas del acrosoma y la superficie del óvulo. De hecho, se habla de reacciones entre «fertilisinas» y «antifertilisinas», con una especificidad comparable a la del encuentro entre enzimas y sustrato.
La singularidad de la fecundación está garantizada por una modificación de la estructura superficial del óvulo («reacción cortical») que comienza justo en el momento de la primera reacción específica de fecundación / antifertilisina; después de esta reacción, la membrana del óvulo se altera, por lo que cualquier otro espermatozoide que lo alcance ya no podrá iniciar la reacción de fecundación específica.
Después de la fecundación, la cola del espermotozoo permanece externa al óvulo, mientras que el material cromosómico lo penetra, este, llamado "pronúcleo masculino", se une al "pronúcleo femenino" del óvulo formando así el núcleo diploide del cigoto.