«Primeros pasos de la glucólisis
La primera enzima utilizada en la segunda fase de la glucólisis es la gliceraldehído 3-fosfato deshidrogenasa; Las deshidrogenasas son enzimas que catalizan la transferencia de poder reductor de una molécula reductora que se oxida a otra molécula que se reduce (reacción redox). Los sustratos de esta enzima son NAD (dinucleótido de nicotidamida y adenina) y FAD (dinucleótido de flavina y adenina).
En este paso, la deshidrogenasa cataliza la conversión de gliceraldehído 3-fosfato en 1,3-bisfosfoglicerato: en el mismo sitio catalítico, el grupo aldehído se oxida a carboxilo con la consiguiente reducción de NAD + a NADH y, posteriormente, el grupo carboxilo es capaz de para formar un enlace anhídrido con un ortofosfato. El primer proceso es muy exergónico (libera energía) mientras que el segundo es muy exergónico (requiere energía); si no hubiera un sitio catalítico, la reacción global no se produciría: la primera reacción se produciría con la liberación de energía que se dispersaría en forma de calor y que, por tanto, no sería utilizable para formar el enlace anhídrido.

Después de la formación de 1,3-bisfosfoglicerato, la enzima reanuda su estructura inicial y está lista para actuar sobre un nuevo sustrato.

Luego viene el fosfoglicerato quinasa que permite la transferencia de un fosforilo de 1,3-bisfosfoglicerato a una molécula de ADP; hemos obtenido ATP (un ATP por cada molécula de gliceraldehído 3-fosfato, por lo tanto, dos ATP por cada molécula de glucosa inicial) que compensa el gasto energético de la primera fase de la glucólisis.
El anión arsenato (AsO43-) afecta la vía glucolítica ya que puede reemplazar al fosfato en la primera reacción de la segunda fase de la glucólisis, dando 1-arsenio 3-fosfoglicerato que es altamente inestable y, tan pronto como está libre del catalizador sitio, hidroliza liberando el "arseniato que regresa a la circulación". Por lo tanto, el arseniato imita la acción del fosfato y entra en el sitio catalítico: en presencia del arsenato, la reacción que produce ATP (de 1,3-bisfosfoglicerato a 3-fosfoglicerato) no tiene lugar porque el 3-fosfato gliceraldehído se convierte directamente en 3-fosfoglicerato; sin ATP disponible, las células mueren (intoxicación por ácido arsénico).
En la tercera reacción de la fase oxidativa, el 3-fosfoglicerato se convierte en 2-fosfoglicerato por la acción de fosfoglicerato mutasa; la reacción implica un intermedio de 2,3-bisfosfoglicerato.

En el siguiente paso, interviene una enzima. enolasa que es capaz de catalizar la eliminación de una molécula de agua del esqueleto carbonoso del 2-fosfoglicerato, obteniendo el fosfoenol pirivado (PEP);

La PEP tiene un alto potencial para transferir un fosforilo: transfiere, a través de la acción de una enzima. piruvato quinasa, un fosforilo a un ADP para dar ATP, en el quinto paso de la segunda fase, obteniendo piruvato.

El 2-fosfoglicerato y el 3-fosfoglicerato tienen un bajo poder de transferencia de un fosforilo, por lo tanto, para obtener ATP a partir de estas moléculas, el 3-fosfoglicerato se convierte en 2-fosfoglicerato, durante la glucólisis, porque se obtiene a partir de este último. especies con alto potencial de transferencia.
Antes de continuar, abramos un paréntesis sobre 2,3-bisfosfoglicerato; este último está presente en todas las células en las que la glucólisis se produce en una concentración muy baja (es el intermedio de la tercera reacción de la segunda fase de la glucólisis). En los eritrocitos, por el contrario, el 2,3-bisfosfoglicerato tiene una concentración estacionaria de 4-5 mM (concentración máxima) porque poseen un patrimonio enzimático que tiene la tarea de producirlo; en los eritrocitos hay una desviación de la glucólisis para producir 2,3-bisfosfoglicerato: el 1,3-bisfosfoglicerato se convierte en 2,3-bisfosfoglicerato por la acción de bisfosfoglicerato mutasa (eritrocito) y 2,3-bisfosfoglicerato, por la acción de bisfosfoglicerato fosfatasa (eritrocito) se convierte en 3-fosfoglicerato. Luego, en los eritrocitos, una parte del 1,3-bisfosfoglicerato obtenido de la glucólisis se convierte en 2,3-bisfosfoglicerato que luego vuelve a la vía glucolítica como 3-fosfoglicerato; al hacerlo, el tercer paso de la fase oxidativa de La glucólisis de la que se obtiene ATP La cantidad de ATP perdido es el precio que un eritrocito está dispuesto a pagar para mantener la concentración de 2,3-bisfosfoglicerato que estas células necesitan porque afecta la capacidad de la "hemoglobina para unirse" al oxígeno.
Hemos visto que en la primera reacción de la segunda fase de glucólisis el NAD + se reduce a NADH pero es necesario que, tras obtener el piruvato, el NADH se reconvierta en NAD +: esto ocurre con la fermentación láctica (se obtiene lactato) o por fermentación alcohólica (entran en juego piruvato descarboxilasa que descarboxila el piruvato y una deshidrogenasa que forma etanol); las fermentaciones no involucran oxígeno (anaerobios).
Debido a la fermentación láctica, el ácido láctico, si no se elimina adecuadamente, se acumula en los músculos y, liberando H +, provoca una contracción muscular involuntaria y, por tanto, calambres; un músculo en fuerte estrés también puede alcanzar un pH mínimo de 6,8.
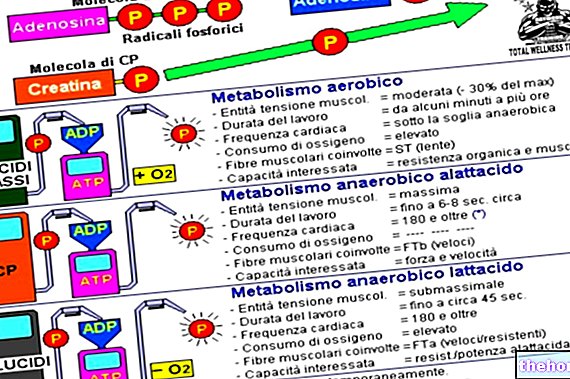
A través del ciclo de Cori, parte de la fatiga de un músculo se transfiere al hígado cuando el músculo está sobrecargado. Suponga que el músculo funciona sin suministro de oxígeno (suposición errónea): si el músculo trabaja moderadamente, el ATP necesario para la contracción lo proporciona exclusivamente la glucólisis. Si la actividad del músculo aumenta y se requiere ATP adicional, acelere el metabolismo aeróbico, convirtiendo lactato, que así se elimina, en glucosa. En realidad, el músculo explota el metabolismo aeróbico: si hay disponibilidad de oxígeno, el músculo explota, sobre todo, el ATP que aporta el metabolismo aeróbico y, cuando no hay más oxígeno disponible, el metabolismo anaeróbico se acelera a través del ciclo de Cori. .Este ciclo asume que el lactato se transfiere del músculo al hígado, donde, al gastar energía, se produce más glucosa que regresa al músculo.A través de este ciclo, parte del ATP consumido en el músculo es suministrado por el hígado que, a través del proceso de gluconeogénesis, es capaz de producir glucosa que puede ser utilizada por el músculo para obtener ATP.
El metabolismo de la glucosa descrito hasta ahora no incluye oxígeno pero el metabolismo aeróbico de la glucosa permite obtener cantidades de ATP 17-18 veces superiores a la obtenida con la vía glucolítica, por lo tanto, cuando la célula tiene la posibilidad de elegir entre aeróbica y ed. anaerobio, favorece al primero.
En el metabolismo aeróbico, el piruvato ingresa a la mitocondria donde sufre transformaciones y eventualmente se obtiene dióxido de carbono y agua; de esta forma se obtienen 34 moléculas de ATP por cada molécula de glucosa degradada.